
Intenzivní trénink aktivuje syntézu bílkovin, ale pouze pokud je k dispozici správná výživa, která ji podporuje. Pokud jste si přečetli článek, který jsme s Johnem Meadowsem napsali Maximized Protein Synthesis, nebo pokud jste četli cokoli, co T Nation na toto téma říká už roky, pravděpodobně jste obeznámeni s pojmem „anabolické okno“ a důležitostí peri-tréninkové výživy.
To, co se děje na buněčné úrovni v hodinách po tréninku, předpovídá dlouhodobé zisky. Využijte tohoto „anabolického okna“ a budete růst jako nikdy předtím. Důsledně chybí a hodně štěstí.
Klíčem je získání správných makroživin ve správný čas, ale makra jsou jen částí celkového obrazu. Důležitým, ale často přehlíženým aspektem syntézy svalových bílkovin je objem buněk. Objem buněk není jen kosmetický; je hlavní hnací silou transportu aminokyselin a také pracuje v zákulisí, aby zapnul syntézu proteinů a potlačil degradaci proteinů.
Plný / objemový sval je anabolický sval. I když již více než 20 let víme, že otoky buněk inhibují štěpení bílkovin a stimulují syntézu bílkovin v určitých buňkách (1-3), donedávna byl mechanismus spojující objem buněk se syntézou bílkovin záhadou.
Nyní víme, že syntéza bílkovin je řízena enzymem mTOR, který je aktivován mechanickým stresem, růstovými faktory a leucinem.
Zatímco všechny tři z nich jsou důležité pro tréninkový stimul, signalizace mTOR je také závislá na objemu buněk.(4) To je zvláště důležité v kosterním svalu, kde volumizace buněk aktivuje syntézu glykogenu a bílkovin a inhibuje rozpad bílkovin.(5, 6)
Vědecký průlom vedoucí ke spojení mezi objemem buněk a syntézou bílkovin nastal v roce 2005, kdy skupina vědců zjistila, že aktivace mTOR vyžaduje více než jen leucin - je také zapotřebí glutamin.(7)
To bylo překvapením. Ačkoli je glutamin považován za „podmíněně esenciální“ aminokyselinu, která omezuje odbourávání bílkovin během těžkého traumatu nebo stresu, nikdy nebyl spojen s aktivací mTOR.
Glutamin byl nezbytný pro absorpci leucinů a objem buněk, což je obojí potřebné k zapnutí syntézy proteinů. Autoři dále ukázali, že buněčný glutamin vyčerpání vede nejen ke snížení objemu buněk, ale také snižuje schopnost leucinů aktivovat syntézu proteinů.(7)
Tento objev byl obrovský, protože poskytoval přímé spojení mezi glutaminem, objemem buněk a syntézou bílkovin. Poprvé se ukázalo, že glutamin je nezbytný pro aktivaci syntézy bílkovin leucinem.
I když tato studie naznačuje, že glutamin je velmi důležitou součástí skládačky spojující objem buněk a syntézu bílkovin, přesný mechanismus nebyl vypracován až do roku 2009, kdy Nicklin et al. zjistili, že export glutaminu je spojen s importem leucinů a aktivací mTOR.(8)
Chcete-li dostat leucin do buňky, je zde počáteční období „glutaminového zatížení."To také přitahuje vodu a zvyšuje objem buněk.". Po fázi „načítání glutaminu“ je glutamin exportován z buňky výměnou za dovoz leucinů.
Nicklin a kol. také objevili, že hladiny buněčného glutaminu omezují rychlost pro aktivaci syntézy proteinů leucinem. Když byly buňky současně ošetřeny glutaminem a směsí EAA obsahující leucin, aktivace syntézy proteinů byla odložena o 60 minut. Když byly stejné buňky „předem naplněny“ glutaminem, byla během 1-2 minut po podání leucinů zapnuta syntéza proteinů.
Tento výsledek byl důležitý, protože v tomto experimentálním modelu vysvětlil dobu zpoždění pro aktivaci syntézy proteinů leucinem.
Tyto výsledky nakonec vrhnou světlo na buněčný aparát, který reguluje transport aminokyselin, a na to, jak je spojen s kontrolou syntézy proteinů.
Tuto práci je však třeba interpretovat s určitou opatrností. Důležitým upozorněním na tyto studie bylo, že byly prováděny in vitro (tj.E. v buněčných kulturách), kde je regulace nebo syntéza proteinů mnohem jednodušší. Svalové buňky jsou schopné produkovat glutamin podle potřeby z jiných aminokyselin a „vyčerpání glutaminu“ v tomto modelu buněčné kultury nepředstavuje fyziologičtější situace in vivo.
Vysoké rychlosti syntézy bílkovin nelze udržovat donekonečna ve svalové tkáni s doplňkem glutaminu nebo bez něj. Glutamin lze však použít ke strategické podpoře syntézy bílkovin optimalizací objemové kapacity buněk během období po cvičení.

Buňky jsou velmi zaneprázdněné a existuje mnoho iontových kanálů a transportních proteinů vázaných na membránu, které regulují přenos do a ven z buňky. Zde jsou důležité zejména dvě třídy transportérů aminokyselin: transportéry aminokyselin „Systém L“ a „Systém A“ jsou nejtěsněji spojeny se signalizací mTOR a syntézou proteinů.(8-10)
Aktivita transportérů systému A a systému L je spojena, což umožňuje absorbovat leucin a další BCAA do buňky.(11) Transportéry systému L jsou odpovědné za příliv leucinů a dalších BCAA výměnou za odtok glutaminu.
Transportéry systému A fungují jiným mechanismem, kde je glutamin spojen s absorpcí sodíku.(12, 13) Vazba mezi příjmem sodíku a transportéry aminokyselin System L / System A se nazývá terciární aktivní transport (zkráceně TAT). Je to TAT, který nakonec pohání leucin uvnitř buňky, což vede k aktivaci mTOR a syntéze bílkovin.(11)
Jak TAT funguje, můžete vidět na obrázku níže:
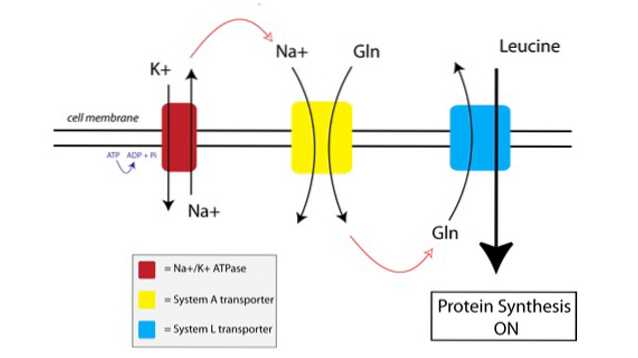
Nejprve membránově vázané čerpadlo nazývané sodno-draselné čerpadlo ATPázy (Na + / K + ATPáza, červené na obrázku výše) využívá energii z ATP k pohybu sodíku mimo buňku proti jeho koncentračnímu gradientu.
Zvýšená koncentrace sodíku mimo buňku je spojena s dovozem glutaminu transportérem systému A (žlutá na obrázku). Přítok glutaminu a sodíku do buňky také přitahuje další vodu, což způsobuje bobtnání buňky. Tím se buňka uvede do anabolického stavu a aktivuje se mechanismus syntézy proteinů.
Když se glutamin hromadí na dostatečně vysoké úrovni uvnitř buňky, aktivují se transportéry systému L (modrý na obrázku), které přepravují glutamin mimo buňku výměnou za vychytávání leucinu. Vstup leucinů do buňky je spouštěčem syntézy proteinů.
I když to byla dosud velká lekce biochemie, objev TAT není důležitý pouze pro buněčné biologové. Nyní, když víme, jak je objem buněk spojen s transportem aminokyselin a syntézou bílkovin, můžeme navrhnout několik výživových strategií, abychom maximalizovali proces, když se to počítá - během kritického období po tréninku.
Syntéza bílkovin je zcela závislá na hydrataci buněk - pokud jste dokonce trochu dehydratovaní, schopnost zotavit se z intenzivního tréninku je zcela narušena. Dostatek vody zde není samozřejmostí, ale samotná voda nestačí.
Elektrolyty, jako je sodík, draslík, chlorid a fosfát, také fungují jako „osmolyty“, protože čerpají vodu do buňky. Po intenzivním tréninku potřebujeme vodu, aminokyseliny a elektrolyty, abychom maximalizovali proces volumizace buněk, který řídí syntézu proteinů.
Zde jsou důležité sodík, hořčík, vápník, draslík, fosfát a chlorid (abychom jmenovali alespoň některé). Na minimální úrovni (pokud tak neučiníte podle doporučení svého lékaře) se nevyhýbejte sodíku před nebo po tréninku. Pokud máte nedostatek sodíku, vaše pumpa z tréninku bude téměř neexistovat a sodík je nezbytný pro absorpci glutaminu.
Abychom eliminovali dohady, jsou Surge® Workout Fuel a Plazma ™ navrženy s ideálními poměry elektrolytů na podporu objemu buněk a syntézy proteinů.
Příjem glutaminu do buňky způsobuje volumizaci buněk a aktivuje svalové buňky pro syntézu proteinů. Jak již bylo zmíněno, plný / objemový sval je anabolický sval. Spolu s řízením transportu aminokyselin zvyšuje objem buněk také syntézu glykogenu a inhibuje rozklad proteinů.(4-6)
Syntéza bílkovin je potlačena deplecí glutaminu, což má obrovské důsledky pro tvrdě trénující sportovce. Po intenzivním tréninku se objeví zánětlivá reakce, která umožňuje imunitním buňkám přenášet se do rozmlácené svalové tkáně a zahájit proces opravy / přestavby.(14)
Glutamin je imunitními buňkami tak rychle přijímán, že je považován za „palivo imunitního systému.„(15) Není divu, že se ukázalo, že intenzivní trénink způsobuje vyčerpání glutaminu v plazmě.(16-18)
Z tohoto důvodu se požadavky na glutamin zvyšují v období po tréninku, kdy místní imunitní odpověď může soutěžit o dostupnost glutaminu pro primární svalové buňky pro příjem aminokyselin a syntézu proteinů.
Předběžné načítání buňky s glutaminem mohou také potenciálně snižovat „zpoždění“ spojené s aktivací proteinu leucinem. Pokud to ještě neděláte, ihned po tréninku užijte 10–15 g glutaminu nebo glutaminových peptidů. Protože BCAA jsou dalším oblíbeným substrátem pro syntézu svalového glutaminu a bylo také prokázáno, že zvyšují produkci svalového glutaminu (19–21), jsou BCAA a leucin také užitečné během období před tréninkem, aby pomohly maximalizovat endogenní produkci glutaminu.
Poznámka redakce: Celý protokol Plamza / MAG-10 s předtréninkovou, tréninkovou a posttréningovou výživou splňuje doporučení autora pro načtení svalů BCAA a leucinem.
Nedávno bylo zjištěno, že spotřeba EAA zvyšuje expresi jak transportérů aminokyselin System A, tak System L.(9) Důležité je, že k tomu dochází na „post-transkripční úrovni“, což znamená na úrovni syntézy proteinů, kde se stávající mRNA překládají na proteiny.
Porovnejte to s „denovo„Exprese proteinu - kde syntéza, zpracování a transport nových mRNA může trvat 16 a více hodin - k posttranskripční aktivaci syntézy proteinů může dojít během několika minut až hodin, což buňkám umožňuje v případě potřeby rychle zvýšit hladinu konkrétních proteinů.
Najednou máme větší podnět k zavedení pevného výživového plánu pro peri-trénink - příjem EAA během období před a peri tréninkem se vyplatí velké po tréninku zvýšením exprese transportérů aminokyselin a aktivací buňky pro maximální příjem aminokyselin a aktivace syntézy proteinů.
Ideální jsou rychle se vstřebávající proteinové izoláty nebo hydrolyzáty, jako je Mag-10® Protein Pulsing Protocol ™ nebo Plazma ™ během období před a po tréninku.
Inzulin je nejvíce anabolický hormon v těle. Spolu s přímou aktivací syntézy proteinů zvyšuje inzulin také translokaci transportérů aminokyselin systému A do buněčné membrány.(22)
To znamená, že inzulín způsobuje, že se na buněčné membráně zobrazí více transportérů systému A, připravených vnést do buňky více glutaminu. Více glutaminu vede k většímu objemu buněk, který do buňky přivádí více leucinu, což nakonec vede k větší syntéze bílkovin.
Zatímco EAA zvyšují výraz transportérů AA, je to inzulinový signál, který jim umožňuje zobrazit se na povrchu buňky, připravený k přenosu nových aminokyselin do buňky.
To je další důvod, proč jsou pre-a peri tréninkové sacharidy dobrým nápadem, pokud nejste v režimu extrémního odbourávání tuků: inzulín zvyšuje kapacitu buněčného transportu aminokyselin.
Sacharidy zvyšují hladinu inzulínu, ale určité aminokyseliny lze také použít k potenciaci uvolňování inzulínu. Glutamin je silný aktivátor „inkretinových“ hormonů, díky nimž jsou buňky produkující inzulín v pankreatu citlivější na glukózu.(23) Glycin také potencuje uvolňování inzulínu jiným mechanismem.
I když samotné sacharidy po tréninku zvýší hladinu inzulínu, kombinace těchto aminokyselin potencujících inzulín s sacharidy přeplní vaši slinivku břišní pro ještě větší uvolňování inzulínu. I když je dobré udržovat hladinu inzulínu na spodní straně po většinu času, zvýšená hladina inzulínu v peri-tréninkovém období maximalizuje transport aminokyselin, objem buněk a syntézu bílkovin a zároveň potlačuje rozklad bílkovin.
Typ intenzivního anaerobního tréninku, který je zapotřebí k vybudování spousty kvalitních svalů, vede ke značné produkci laktátu a ke snížení svalového pH. To vede k časné svalové únavě a slabosti, ale také určité transportéry aminokyselin, včetně systému A, jsou inhibovány nízkým pH.(13)
Když je pH svalů nízké, absorpce aminokyselin je snížena, což potlačuje aktivaci syntézy proteinů mTOR.(24) Rovněž se ukázalo, že nízká inhibice pH transportérů aminokyselin systému A zvyšuje rozklad bílkovin.(25)
To je místo, kde přichází beta-alanin. Zvýšené hladiny karnosinu ve svalech fungují jako přírodní kyselý pufr, čímž se prodlužuje anaerobní prahová hodnota omezením poklesu svalového pH při tréninku.
Beta-alanin má další důležitou funkci: pomáhat udržovat syntézu bílkovin a dostat ji rychleji online po intenzivním tréninku prevencí útlumu transportu aminokyselin.
Chcete-li zvýšit hladinu svalového karnosinu, užívejte dvě tablety Beta-7 ™ třikrát denně.
Během intenzivních tréninků se snižuje syntéza bílkovin a aktivuje se degradace bílkovin. To je nevyhnutelné pro každého tvrdě trénujícího zvedače. Avšak to, do jaké míry můžeme minimalizovat katabolické účinky tréninku a čím rychleji se můžeme vrátit do „anabolického režimu“ během období po tréninku, nakonec určuje, jak efektivně se zotavíme - a budeme růst.
Načasování makroživin je důležité, ale je to prostředek k dosažení cíle. Objem buněk je hlavní hnací silou přenosu aminokyselin a syntézy bílkovin. Pochopením toho, jak dochází k transportu aminokyselin a jak je regulován objemem buněk, můžeme rychleji dostat více leucinu do zničených svalových buněk, čímž podnítíme anabolický oheň a nakonec to povede k lepším ziskům.
Výše uvedené strategie jsou účinné, praktické a jsou založeny na nejnovějších vědeckých výzkumech. Použijte je jako šablonu a posuňte svůj tréninkový pokrok na další úroveň.
Těším se na vaše dotazy týkající se LiveSpill!


Zatím žádné komentáře